Adrenalina y Noradrenalina.
agosto 30, 2007 at 12:13 pm 8 comentarios
El primer lugar de síntesis de adrenalina es en la médula suprarrenal, partir de al cual se libera directamente sobre el torrente sanguíneo, la síntesis es llevada a cabo por metilación de al noradrenalina mediante al enzima adrenalina n-metiltransferasa utilizando la s-adenosilmetionina como cofactor. La liberación se da por despolarización por el potasio y por otros tratamientos despolarizantes, este mecanismo es dependiente de calcio. El transporte de alta afinidad de la adrenalina hasta los terminales nerviosos y células gliales, es casi con certeza el método principal mediante el cual se inactiva la adrenalina liberada en las sinapsis. Aun no se ha logrado desarrollar un fármaco que posee una especificidad adecuada con respecto a los sistemas adrenérgicos.


La adrenalina esta involucrada en:
- Mecanismos centrales de control vasomotor y respiración
- Termoregulación
- Regulación de al ingesta de alimentos y agua
- Control de la secreción pituitaria

 Aunque la adrenalina puede funcionar como neurotransmisor, su papel en el funcionamiento del SNC queda en realidad completamente relegado por la acción de la noradrenalina; si bien utilizamos generalmente el termino adrenérgico. Esta paradoja se debe a que la potente producción de adrenalina desde la médula de las glándulas suprarrenales, como consecuencia de la activación simpática, tiene unas consecuencias generalizadas e iguales que las de la acción de la noradrenalina liberada por la neurona postsináptica de una vía autónoma.
Aunque la adrenalina puede funcionar como neurotransmisor, su papel en el funcionamiento del SNC queda en realidad completamente relegado por la acción de la noradrenalina; si bien utilizamos generalmente el termino adrenérgico. Esta paradoja se debe a que la potente producción de adrenalina desde la médula de las glándulas suprarrenales, como consecuencia de la activación simpática, tiene unas consecuencias generalizadas e iguales que las de la acción de la noradrenalina liberada por la neurona postsináptica de una vía autónoma.
La noradrenalina es, por tanto, la catecolamina que se utiliza como neurotransmisor en el sistema nervioso central (SNC), y podemos decir que la masa más compacta y densa de neuronas adrenérgicas la constituye el locus ceruleus, el cual está perfectamente identificado en el tronco cerebral.
Precisamente es desde el locus ceruleus, y también desde otras áreas noradrenérgicas inferiores como el núcleo del tracto solitario o los núcleos reticulares laterales, desde donde surgen dos grandes fascículos de proyección ascendente: el fascículo noradrenérgico dorsal y el fascículo noradrenérgico ventral. Ambos fascículos tienen una especial implicación en el nivel de actividad córtico-subcortical precisamente por su proyección al córtex, al sistema límbico (hipocampo, amígdala y septum) y al diencéfalo, tálamo e hipotálamo. Asimismo, se proyectan descendentemente a la formación reticular de la médula, con lo que su principal papel estriba en la regulación de los niveles de vigilancia y, particularmente, en la actividad mínima de la vigilia, definiendo claramente los niveles de atención, emoción e hiperexcitabilidad; por lo que su repercusión conductual es tremendamente significativa, si bien es cierto que la división simpática del sistema nervioso autónomo también tiene una especial relevancia.
Metabolismo y distribución
 La dopa decarboxilasa es un enzima piridoxinodependiente que cataliza la eliminación del grupo carboxilo de la dopa para formar dopamina. La dopa decarboxilasa puede también decarboxilar el 5-hidroxitriptófano, precursor de la serotonina, así como otros aminoácidos aromáticos. La dopa decarboxilasa se encuentra ampliamente distribuida por todo el cuerpo, donde se puede hallar tanto en neuronas que contienen catecolaminas como en aquellas que contienen serotonina, y en tejidos no neuronales, tales como el riñón o los vasos sanguíneos. La a-metildopa inhibe la DDC in vitro, e induce a una reducción en la presión sanguínea, tras ser convertida en el falso transmisor a-metil-norepinefrina.
La dopa decarboxilasa es un enzima piridoxinodependiente que cataliza la eliminación del grupo carboxilo de la dopa para formar dopamina. La dopa decarboxilasa puede también decarboxilar el 5-hidroxitriptófano, precursor de la serotonina, así como otros aminoácidos aromáticos. La dopa decarboxilasa se encuentra ampliamente distribuida por todo el cuerpo, donde se puede hallar tanto en neuronas que contienen catecolaminas como en aquellas que contienen serotonina, y en tejidos no neuronales, tales como el riñón o los vasos sanguíneos. La a-metildopa inhibe la DDC in vitro, e induce a una reducción en la presión sanguínea, tras ser convertida en el falso transmisor a-metil-norepinefrina.
Para las neuronas que sintetizan adrenalina o noradrenalina, la dopamina-b-hidroxilasa es el siguiente paso en la ruta biosintética.
Al igual que la TH, la dopamina-b-hidroxilasa (DBH) es una oxidasa de función combinada que usa el oxígeno molecular para formar el grupo hidroxilo añadido al b-carbono en la cadena lateral de la dopamina. La dopamina-b-hidroxilasa contiene Ca+2, el cual está impolicado en la transferencia de electrones en la reacción; así, los quelatos de cobre tales como el dietilditiocarbonato son potentes inhibidores de la DBH.
 En las células cromafínicas que sintetizan adrenalina, el paso final de la ruta está catalizado por el enzima feniletanolamina N-metiltransferasa.
En las células cromafínicas que sintetizan adrenalina, el paso final de la ruta está catalizado por el enzima feniletanolamina N-metiltransferasa.
La feniletanolamina N-metiltransferasa (PNMT) transfiere un grupo metilo desde la S-adenosilmetionina al nitrógeno de la noradrenalina, formando una amina secundaria. La actividad de la PNMT está regulada por los corticoesteroides. La alta actividad de la PNMT en la médula adrenal refleja las altas concentraciones de corticoesteroides liberados dentro de los senos venosos que irrigan el cortex adrenal. La hipofisectomía, que causa una disminución en el nivel de corticoesteroides, conduce a marcadas reducciones en la cantidad de este enzima; inversamente, la administración de grandes cantidades de corticoesteroides, conlleva a la síntesis de PNMT en las neuronas simpáticas.
 Generalmente encontramos catecolaminas en una baja concentración de forma libre en el citosol, donde pueden ser metabolizadas por diversos enzimas incluyendo las monoaminooxidasas (MAO). De esta forma la conversión de tirosina a L-dopa, y de L-dopa a dopamina, tiene lugar en el citosol; tras esto, la dopamina es recogida por las vesículas de almacenamiento.
Generalmente encontramos catecolaminas en una baja concentración de forma libre en el citosol, donde pueden ser metabolizadas por diversos enzimas incluyendo las monoaminooxidasas (MAO). De esta forma la conversión de tirosina a L-dopa, y de L-dopa a dopamina, tiene lugar en el citosol; tras esto, la dopamina es recogida por las vesículas de almacenamiento.
La b-hidroxilación final se produce en el interior de estas vesículas. El mecanismo que concentra a las catecolaminas en el interior de las vesículas es un proceso dependiente del adenosín-trifosfato (ATP) ligado a una bomba de protones. El proceso de recaptación vesicular tiene una amplia especificidad de substrato y es capaz de transportar varias aminas biogénicas, incluyendo a la triptamina, la tiramina y a las anfetaminas; estas aminas pueden competir con las catecolaminas endógenas por ocupar un lugar en las vesículas de almacenamiento. La reserpina es un inhibidor específico e irreversible de la bomba de aminas vesicular que acaba con la capacidad de estas vesículas para concentrar las aminas. El tratamiento con reserpina provoca una profunda reducción de catecolaminas endógenas en las neuronas. El efecto de la reserpina es el de inhibir la recaptación de dopamina y otras catecolaminas en el interior de las vesículas.
Cuando un potencial de acción alcanza el terminal nervioso, se abren los canales de Ca+2, permitiendo un influjo del catión en el terminal; el incremento del Ca+2 intracelular promueve la fusión de las vesículas con la membrana neuronal. Entonces las vesículas descargan sus contenidos solubles, incluyendo noradrenalina, ATP y DBH dentro del espacio extraneuronal. La liberación exocitótica desde las neuronas simpáticas podría ser el orígen de algunas de las DBH que se han encontrado en el plasma y en el fluido cerebroespinal de animales y humanos.
Simpaticomiméticos de acción indirecta, como la tiramina y la anfetamina, liberan catecolaminas por un mecanismo que ni depende del Ca+2 ni se encuentra asociado con la liberación de DBH.
Hay mecanismos reguladores que operan eficientemente para modular la proporción de síntesis de catecolaminas. Un proceso de larga duración que afecte a la síntesis de catecolaminas trae consigo alteraciones en las concentraciones de TH y DBH presentes en los terminales nerviosos. Cuando el nivel de actividad neuronal de las neuronas simpáticas es incrementado durante un largo periodo de tiempo, las concentraciones de ARNm que codifican la TH y la DBH se incrementan en la pericarión neuronal.
 Dos son los enzimas principalmente responsables de la inactivación de las catecolaminas: las monoamino oxidasas y la catecol-Oxi-metiltransferasa.
Dos son los enzimas principalmente responsables de la inactivación de las catecolaminas: las monoamino oxidasas y la catecol-Oxi-metiltransferasa.
Las monoamino oxidasas y la catecol-Oxi-metiltransferasa (COMT) se encuentran ampliamente distribuidas por todo el cuerpo. La monoamino oxidasa es un enzima que contiene flavina localizada sobre la membrana exterior de la mitocondria. Este enzima deaminado oxidativamente las catecolaminas a sus correspondientes aldehídos; estos pueden ser convertidos, alternativamente, por aldehído deshidrogenasa en ácidos, o por aldehído reductasa para formar glicoles. Debido a su localización intracelular, la MAO desmpeña un papel estratégico en la inactivación de las catecolaminas que se encuentran líbremente dentro del terminal nervioso y no están protegidas por las vesículas de almacenamiento. De acuerdo con esto, las drogas que interfieran con el almacenamiento vesicular, como la reserpina, anfetaminas, que desplazan a las catecolaminas de las vesículas, provocan un incremento significativo en los metabolitos deaminados. La MAO-A preferentemente deamina a la noradrenalina y a la serotonina, y es selectivamente inhibida por la clorgilina, mientras que la MAO-B actúa sobre un amplio espectro de feniletilaminas, entre las que se incluye la b-feniletilamina. La monoamino oxidasa B es selectivamente inhibida por el deprenil. La monoamino oxidasa desmpeña una importante función protectora en el tracto gastrointestinal y en el higado, previniendo el acceso de las aminas de acción indirecta que se encuentran en la comida, tales como la tiramina y la feniletilamina, a la circulación general; de esta forma los pacientes tratados de depresión o hipertensión con inhibidores de la MAO no gozarían de esta protección y podrían sufrir graves crisis hipertensivas tras la ingestión de alimentos que contengan grandes cantidades de tiramina, alimentos como el queso, los arenques y el vino de Oporto. Si sustituímos por un metilo el a-carbono de la cadena lateral de la feniletilamina, éste protegerá contra la deaminación por la MAO; la acción prolongada de las anfetaminas y otros estimulantes de acción indirecta es en parte una consecuencia de la presencia de un grupo a-metilo, el cual previene su inactivación por la MAO.
 La catecol-O-metiltransferasa se encuentra en casi todas las células, entre las que se incluyen a los eritrocitos. De esta forma, la COMT actúa sobre las catecolaminas extraneuronales. El enzima, que requiere Mg+2, transfiere un grupo metilo desde el cosubstrato S-adenosilmetionina al grupo 3-hidroxi en el anillo de la catecolamina.
La catecol-O-metiltransferasa se encuentra en casi todas las células, entre las que se incluyen a los eritrocitos. De esta forma, la COMT actúa sobre las catecolaminas extraneuronales. El enzima, que requiere Mg+2, transfiere un grupo metilo desde el cosubstrato S-adenosilmetionina al grupo 3-hidroxi en el anillo de la catecolamina.
El ácido 4-hidroxi-3-metoxi-fenilacético, más conocido comúnmente como ácido homovanílico (HVA), es un metabolito principal de la dopamina. Los niveles de este ácido en el líquido cefalorraquídeo (LCR) proporcionan una idea acerca de la renovación de la dopamina en el estriado. Los niveles de HVA disminuyen, por ejemplo, en el fluido cerebroespinal de pacientes con la enfermedad de Alzheimer. Un metabolito de la noradrenalina formado de manera relativamente selectiva en el cerebro es el 3-metoxi-4-hidroxi-fenilglicol (MHPG). Se estima que entre un 30 y un 50% del MHPG excretado en la orina se deriva del cerebro.
El proceso de recaptación de catecolaminas fue originalmente descrito por Axelrod, y depende de la energía, ya que puede ser inhibido por incubación a baja temperatura o por inhibidores metabólicos. Los requerimientos energéticos reflejan un acoplamiento del proceso de captación con el gradiente de Na+ a lo largo de la membrana neuronal; algunas drogas que inhiben la Na, K-ATPasa, u otras como la veratridina, que abre los canales de Na+ puede tener significancia fisiológica ya que el transporte cesa al mismo tiempo que la liberación de catecolaminas inducida por despolarización. La captación de catecolaminas puede inhibirse selectivamente por drogas tales como los antidepresivos tricíclicos y la cocaína, además de varias feniletilaminas, tales como las anfetaminas ligadas al portador; de esta forma, pueden estar concentradas dentro de las neuronas que contienen catecolaminas, y pueden competir con las catecolaminas por el transporte. La captación a través de la membrana de la vesícula requiere Mg+2.
Los cuerpos celulares de las neuronas noradrenérgicas están agrupados en la médula oblongada, el puente y el cerebro medio, y se considera que son anatómicamente una parte de la formación reticular.
Las fibras noradrenérgicas pueden dividirse dentro de dos vías fundamentales: el haz dorsal y el ventral. Los cuerpos celulares originados en el haz dorsal se encuentran contenidos en un denso núcleo conocido como locus coeruleus, situado lateralmente sobre el cuarto ventrículo.
La noradrenalina se biosintetiza en las terminaciones sinápticas a partir del aminoácido tirosina por acción de la tirosina hidroxilasa, produciéndose la dopa la cual, mediante la dopa descarboxilasa se convierte en dopamina (DA), la primera de las catecolaminas. La dopamina, por hidroxilación con la b-hidroxi-dopamina se transforma en noradrenalina (NA), que es la segunda de las catecolaminas. Finalmente, la NA, por una metilación con la feniletanolamina N-metiltransferasa (PNMT), se convierte en adrenalina (A).
La diferenciación de las neuronas por su actividad dopaminérgica, noradrenérgica o adrenérgica ha de basarse, en primer lugar, en la existencia de una actividad enzimática tirosina hidroxilasa, y en segundo lugar en la ausencia de dopamina b-hidroxilasa, para las neuronas dopaminérgicas, o en ausencia de actividad PNMT para las neuronas noradrenérgicas. Lógicamente, la presencia de estas actividades enzimáticas caracteriza a la neurona como adrenérgica.
Por lo que respecta a su almacenamiento en el botón terminal, hay una clara diferencia con respecto a las vesículas sinápticas conocidas para la acetilcolina, y es que las catecolaminas se almacenan conjugadas a una proteína ácida, la cromogranina, que protege al neurotransmisor de la acción de las enzimas degradantes; pero que, sin embargo, es fácilmente separable y reparable por la intervención del ATP.
En este proceso es importante el significado funcional de la tirosina hidroxilasa, puesto que por una parte, es el punto de retroalimentación en la homeostasis de las catecolaminas, además de ser fácilmente inhibida por la concentración del propio neurotransmisor, lo que impide su acumulación excesiva. Pero también es el lugar de acción de distintos fármacos o drogas como la a-metil paratirosina, que mediante la inhibición de la tirosina hidroxilasa producen una paulatina y potente deplección en la concentración de noradrenalina cerebral.
 Resulta también muy interesante el hecho de que los distintos sustratos, que procediendo del metabolismo endógeno, pueden producir o bien un incremento de los neurotransmisores o bien, por el contrario, antagonizar su efecto y depleccionar la neurona. Este es el caso de los denominados falsos neurotransmisores, como la octopamina (que se forma por descarboxilación de tirosina a tiramina y su posterior b-hidroxilación), o la a-metil-octopamina, (que se forma en el metabolismo de las anfetaminas) y que tanto por desplazar a la noradrenalina de su conjugación proteica, como por competir con los sistemas enzimáticos de su degradación, provocan en ambos casos un incremento en la acción neurotransmisora con la consecuente complicación fisiológica como la hipertensión. Sin embargo, estas estructuras que bloquean la acción ganglionar, como la reserpina, la guanetidina y el fosfato de bretilio que disminuyen la actividad del neurotransmisor con una clara deplección, pueden ser utilizadas como hipotensores, sedantes y antiarrítmicos.
Resulta también muy interesante el hecho de que los distintos sustratos, que procediendo del metabolismo endógeno, pueden producir o bien un incremento de los neurotransmisores o bien, por el contrario, antagonizar su efecto y depleccionar la neurona. Este es el caso de los denominados falsos neurotransmisores, como la octopamina (que se forma por descarboxilación de tirosina a tiramina y su posterior b-hidroxilación), o la a-metil-octopamina, (que se forma en el metabolismo de las anfetaminas) y que tanto por desplazar a la noradrenalina de su conjugación proteica, como por competir con los sistemas enzimáticos de su degradación, provocan en ambos casos un incremento en la acción neurotransmisora con la consecuente complicación fisiológica como la hipertensión. Sin embargo, estas estructuras que bloquean la acción ganglionar, como la reserpina, la guanetidina y el fosfato de bretilio que disminuyen la actividad del neurotransmisor con una clara deplección, pueden ser utilizadas como hipotensores, sedantes y antiarrítmicos.
 La liberación de NA se debe al flujo de iones Ca2+ que acompañan al potencial de acción, de igual forma que con la acetilcolina, y éste es el camino principalmente utilizado por los estimulantes, como las anfetaminas, para conseguir un mayor efecto de activación postsináptica. Es decir, que sin modificar la actividad de los receptores producen un aumento de la respuesta por facilitación de la liberación de neurotransmisores. También, y secundariamente, las anfetaminas refuerzan su efecto catecolaminérgico por la inhibición de la recaptación presináptica del neurotransmisor.
La liberación de NA se debe al flujo de iones Ca2+ que acompañan al potencial de acción, de igual forma que con la acetilcolina, y éste es el camino principalmente utilizado por los estimulantes, como las anfetaminas, para conseguir un mayor efecto de activación postsináptica. Es decir, que sin modificar la actividad de los receptores producen un aumento de la respuesta por facilitación de la liberación de neurotransmisores. También, y secundariamente, las anfetaminas refuerzan su efecto catecolaminérgico por la inhibición de la recaptación presináptica del neurotransmisor.
Una vez liberada una catecolamina, el principal mecanismo de desactivación es la recaptación. En la recaptación participa activamente la membrana presináptica con un mecanismo similar al de la bomba de Na+, que se denomina bomba de amina. Es en este fenómeno donde los fármacos tricíclicos presentan su actividad inhibiendo la recaptación, por lo que su efecto consiste claramente en una potenciación de las catecolaminas; de ahí se deriva el valor terapéutico de los antidepresivos tricíclicos.
La acción enzimática, aunque minoritaria con respecto a la recaptación, también constituye una parte importante en la inactivación de las catecolaminas. Los sistemas enzimáticos implicados son dos, la monoaminooxidasa (MAO) y catecol-oxi-metil-transferasa (COMT). Estos sistemas actúan de modo sucesivo y oxidan el grupo amino e incorporan un grupo metilo respectivamente. Así, la inhibición específica de los sistemas enzimáticos, principalmente de la MAO, da lugar a un grupo denominado inhibidores de la MAO (IMAOs), como la tranilcipromina, la clorgilina, el deprenilo, la pargilina y otros. Lógicamente, su acción terapéutica es antidepresiva al aumentar la potencia de acción de las catecolaminas. Sin embargo, los efectos secundarios, particularmente la toxicidad hepática y la hipertensión, han relegado su uso aunque actualmente se investiga una nueva generación de IMAOS de corta duración y sobre todo de reversibilidad en su efecto enzimático como la meclobemida, que no presenta hepatotoxicidad y que reconduce el uso clínico de estos antidepresivos.
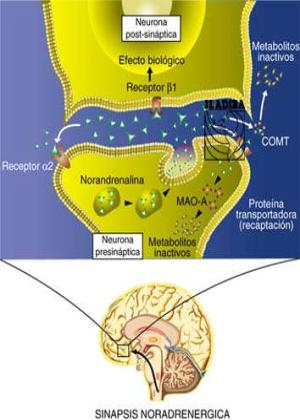
Sinapsis noradregérgica.
La norepinefrina (noradrenalina) activa la neurona postsináptica y después, la COMT (catecoloximetiltransferasa) transforma la norepinefrina en normetanefrina. Además es recaptada por la neurona presináptica. Cuando se libera la Norepinefrina:
Actúa sobre el receptor postsináptico. Actúa sobre el receptor presináptico (ponen en marcha un mecanismo feed-back negativo y frena la liberación de nueva norepinefrina). La COMT degradaría la norepinefrina. La MAO (monoaminooxidasa) rompe la norepinefrina que se libera en las vesículas presinápticas. Cuando se libera hay más norepinefrina. Los inhibidores de MAO son los IMAO.

El torrente circulatorio lleva y hace ingresar a la neurona el aminoácido precursor Tirosina. Una enzima, la Tirosina-hidroxilasa transforma la Tirosina en DOPA. La dopa-descarboxilasa transforma la DOPA en Dopamina. Dentro las vesículas de almacenamiento la Dopamina-B-hidroxilasa transforma a la Dopamina en Noradrenalina*. Al unirse la Noradrenalina con su receptor en el botón postsináptico, provoca cambios en la permeabilidad de la membrana, favoreciendo intercambio más rápido de iones, iniciando un nuevo impulso eléctrico. La acción del neurotransmisor también podría inducir eventos intracelulares, activando segundos mensajeros que podrían activar incluso procesos de replicación proteica a partir del ADN en el núcleo de la neurona. La Catecol-orto-metiltransferasa desactiva a la norepinefrina transformandola en Metoxihidroxifeniglicol para su eliminación.Otro mecanismo de eliminación de la norepionefrina es su recaptura para ser transformada por la Monoaminoxidasa en MHFG.
* Las neuronas dopaminérgicas no tienen la enzima Dopamina-b-hidroxilasa que transforme a la Dopamina en Noradrenalina, por lo tanto usan la Dopamina como neurotransmisor.

Receptores de las catecolaminas



Autorreceptores o prerreptores .
Los autorreceptores que se encuentran en cualquier neurona responden a las moléculas transmisoras liberadas por la misma neurona. En el terminal nervioso, responden a las moléculas transmisoras liberadas en la hendidura sináptica; sobre el cuerpo celular responderían a las moléculas transmisoras liberadas por las dendritas. Funcionalmente, la mayoria de los autorreceptores parecen regular la liberación de transmisores. Se han identificado los autorreceptores para las neuronas que contienen noradrenalina, dopamina, serotonina y GABA. Los principales tipos de autorreceptores inhibidores, descritos tanto en el sistema nervioso simpático periférico como en el cerebro, tienen propiedades farmacológicas similares a aquellas de los receptores ?a-adrenérgicos.
Los autorreceptores del tipo b-adrenérgico también han sido descritos. Esto difiere de la mayoría de los otros autorreceptores conocidos, en los que la noradrenalina que actúa sobre estos receptores facilita la liberación de transmisores, y de esta forma amplifica los efectos del bombardeo neuronal. Este efecto contrasta con la acción inhibidora de los autorreceptores a-adrenérgicos y de la dopamina, los cuales ejercen un control de retroalimentación negativa sobre la liberación de transmisores.
Receptores postsinápticos
Los efectos de la dopamina están mediados a través de la interacción con los receptores del tipo D1 (D1 y D5) y D2 (D2, D3 y D4), mientras que los efectos de la noradrenalina y la adrenalina están mediados a través de los receptores a-adrenérgicos (a1- y a2) y a través de los receptores b-adrenérgicos (b1 y b2).
Tanto la noradrenalina como la adrenalina actúan sobre los receptores a y b, pero el isoproterenol, un agonista sintético, actúa sólo sobre los receptores b. El antagonista receptor b-adrenérgico propanolol está esencialmente inactivo en los receptores a; el antagonista receptor a-adrenérgico pentolamina es muy débil en los receptores b.
De los receptores b, los b1-adrenérgicos, son los que predominan en el corazón y en el córtex cerebral, mientras que los receptores b2-adrenérgicos predominan en el pulmón y en el cerebelo. Sin embargo, en muchos casos los receptores b1- y b2-adrenérgicos coexisten en el mismo tejido. Un importante efecto secundario de los agonistas selectivos b2 como el metaproterenol, usado para tratar el asma broquial, es la aceleración cardíaca.
El cerebro contiene tanto receptores b1 como b2, a los que no se puede diferenciar en virtud de sus funciones fisiológicas.
Se ha identificado un tercer tipo de receptor b-adrenérgico, los receptores b3 en el tejido adiposo marrón presente en roedores y en humanos recién nacidos.
Receptores a-adrenérgicos
Algunos tejidos poseen únicamente receptores postsinápticos a1, otros poseen receptores postsinápticos a2 y algunos órganos tienen una mezcla de los dos. Las consecuencias fisiológicas de los dos tipos de receptores a en el cerebro no están claras actualmente. Es llamativo que la especificidad a los fármacos de los receptores a2 se parezca muchísimo a la de los autorreceptores adrenérgicos, lo cual también hace referencia a los receptores a2.
Es importante conocer los receptores adrenérgicos por el gran significado funcional que depende de la acción de los mismos tanto a nivel central como periférico. Hay dos grandes grupos de receptores adrenérgicos: a y b, que se subdividen a su vez en a1 y a2, y en b1 y b2, respectivamente. Los receptores a1 son postsinápticos y los a2 son presinápticos, de tal manera que es fácil entender el efecto autorregulador de la actividad a2-adrenérgica y el efecto excitatorio de la actividad a1-adrenérgica. Sin embargo, los receptores b1 y b2 son postsinápticos, pero con unos efectos claramente diferenciales; mientras que los b1 actúan sobre la musculatura cardiaca, los b2 actúan sobre la musculatura lisa bronquial y vascular.
Podemos inmediatamente platearnos la dificultad de definir un efecto adrenérgico de una manera general, ya que, por ejemplo, un uso terapéutico adrenérgico que puede estar perfectamente indicado en un caso de broncodilatación, puede ser tremendamente perjudicial para un sujeto por sus efectos cardiacos.
Existen efectos a-adrenérgicos típicos como en el caso de la adrenalina y la noradrenalina, que presentan una afinidad particular por los receptores a1 y a2, de manera que la respuesta fisiológica depende de la regulación de la actividad a1 y a2.
Un agonista a1-adrenérgico, como la fenilefrina, potencia el efecto fisiológico y, por ejemplo, tiene una clara indicación en la hipotensión. Sin embargo, la clonidina, que es un agonista a2-adrenérgico, inhibe con su excitabilidad la liberación de noradrenalina, originando una disminución de la actividad adrenérgica postsináptica, lo que le da un uso terapéutico como claro antihipertensor. Como podemos deducir, estamos ante un mismo mecanismo de manera que, utilizando agonistas adrenérgicos podemos obtener consecuencias totalmente opuestas de acuerdo con la especificidad del receptor sobre el que tienen afinidad.
Esta misma situación debemos plantearnosla con los antagonista a-adrenérgicos. Un antagonista a1 inhibiría o bloquearía el efecto postsináptico de la noradrenalina y, sin embargo, un antagonista a2 aumentaría los efectos postsinapticos al no regular la liberación de noradrenalina por bloqueo de sus autorreceptores.
Receptores b-adrenérgicos
Hemos de significar también los efectos b-adrenérgicos. La isoprenalina o isoproterenol, agonistas b-adrenérgicos, presenta una máxima actividad adrenérgica tanto sobre receptores b1 como b2. Su efecto es el doble que el de la adrenalina, cuatro veces más potente que el de la noradrenalina sobre los b1, y cincuenta veces más potente que el de esta misma sobre los b2, lo que muestra el escasísimo efecto de la noradrenalina sobre los receptores b2-adrenérgicos. Es díficil encontrar un agonista puro para alguno de los receptores b y, por tanto, lo que se observa como parámetro de valoración terapéutica es la relación entre los efectos b2-b1. En este sentido, aquellos agonistas con un mayor efecto b2, como por ejemplo el salbutamol, que es sesenta veces más potente en los receptores b2 que en los b1, está indicado ante la presencia de problemas asmáticos y broncopulmonares, precisamente por su bajísimo efecto cardiovascular.
Los antagonistas b-adrenérgicos o b-bloqueantes constituyen un grupo enormemente representativo por su enorme aplicación antihipertensora, antiarrítmica y también ansiolítica. Entre otros, son antagonistas b-adrenérgicos el propanolol, el pronetalol, el oxprenolol y otros, aunque debemos tener en cuenta que su uso es perjudicial en individuos con problemas asmáticos.
Entry filed under: Documentos de importancia, Neurobiología.
8 comentarios Add your own
Deja un comentario
Trackback this post | Subscribe to the comments via RSS Feed

{kind=link}
1. jose ivan (chile) | septiembre 6, 2007 a las 12:52 am
jose ivan (chile) | septiembre 6, 2007 a las 12:52 am
me parecio muy interesante su web
2. Gustavo David | septiembre 16, 2007 a las 10:19 pm
Gustavo David | septiembre 16, 2007 a las 10:19 pm
Sera cierto que los B3 se encuentran en tejido adiposo, pues decimos que los adrenergicos actuan sobre corazon o pulmon pero a que se refiere con humano recien nacido.
3. fassora marcela | septiembre 22, 2007 a las 8:49 pm
fassora marcela | septiembre 22, 2007 a las 8:49 pm
Mi consulta es referida al tema, tengo inconvenientes para dormir, me levanto fatigada, problemas de hipersensibilidad,miedos, fobia social,tengo 38 años no puedo trabajar como consecuencia de todos estos inconvenientes, e incluso afectaron mi función renal que ya era monorrea,la medicación que me dan no me sienta bien,
me tiene durmiendo todo el tiempo,después de tomarla no me puedo levantar de la cama, incluso me aumenta la presión arterial, ya estoy medicada por el tema renal,la medicación que me dieron fue Quilarex, me gustaria saber que podria tomar de manera que pudiera estabilizar la recaptación de serotonina y noradrenalina, siendo una persona normal, que pueda estar estabilizada de manera que pueda trabajar. Desde ya lo saluda Atte. Quedando a la espera de una respuesta suya.
4. NANCY | enero 21, 2008 a las 8:10 pm
NANCY | enero 21, 2008 a las 8:10 pm
informacion acerca de articulos educativos de la noradrenalina
5. VANESSA | enero 22, 2008 a las 9:34 pm
VANESSA | enero 22, 2008 a las 9:34 pm
A mi me han hecho una analitica especial de orina y me ha dicho la endocrina que los resultados no son muy normales era para ver si me dais vuestra opinion sobre estos resultados metanefrina 49,40 normetamefrina 78,0 diuresis orina con acido 800ml adrenalina 1,32
6. nancy cabas siñani | marzo 6, 2008 a las 9:01 pm
nancy cabas siñani | marzo 6, 2008 a las 9:01 pm
informacion sobre la nor-adrenalina
7. Cory | marzo 25, 2009 a las 2:10 am
Cory | marzo 25, 2009 a las 2:10 am
Muy útil esta información! Gracias desde Venezuela!
8. hector guerra | julio 20, 2009 a las 11:07 pm
hector guerra | julio 20, 2009 a las 11:07 pm
exelent i think that is so much important to know about this topic. pleas send me another report about this