El hidrógeno es un elemento fundamental en los organismos vivos. De hecho, la existencia de la vida depende de dos propiedades especificas del hidrógeno: la cercanía de las electronegatividades del carbono y el hidrógeno y su capacidad para formar puentes cuando esta unido de forma covalente con N y O. La baja polaridad del enlace C-H contribuye a la estabilidad de los compuestos orgánicos en nuestro mundo tan reactivo químicamente. Por ejemplo: los lípidos, las proteínas, en el DNA y RNA.
Un enlace de hidrógeno se forma entre moléculas polares con hidrógeno unido covalentemente a un átomo pequeño muy electronegativo, como flúor, oxígeno o nitrógeno ( F-H, O-H, N-H ).
Un puente de hidrógeno es en realidad una atracción dipolo-dipolo entre moléculas que contienen esos tres tipos de uniones polares.
Los enlaces de hidrógeno tienen solamente una tercera parte de la fuerza de los enlaces covalentes, pero tienen importantes efectos sobre las propiedades de las sustancias en que se presentan, especialmente en cuanto a puntos de fusión y ebullición en estructuras de cristal. Los compuestos con mayor capacidad para formar puentes de hidrógeno son los más electronegativos como N, O y F. Algunos compuestos que poseen enlace hidrógeno son:

Ahora bien, realmente existen dos clases de enlace o puente hidrógeno:
- enlace hidrógeno intermolecular
- enlace hidrógeno intramolecular.
El enlace hidrógeno intermolecular, se realiza entre moléculas, así por ejemplo:

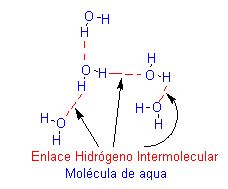
Los puentes de hidrógeno entre moléculas de agua tienen especial importancia para la vida en nuestro planeta. Sin puentes de hidrógeno, el agua se fundiría a unos –100° C y haría ebullición a cerca de –90° C. Los puentes dan pie a otra propiedad muy poco común del agua: la fase liquida es más densa que la fase sólida. Las moléculas de la mayor parte de las sustancias están «apretadas» en la fase sólida que en la líquida, por lo que el sólido es más denso que el líquido.
Por ello, un sólido casi siempre se hunde al fondo cuando comienza a cristalizarse a partir de la fase líquida. Si esto ocurriera con el agua, los lagos, ríos y mares de las regiones en las que las temperaturas bajan de cero, se congelarían desde el fondo hacia arriba. Es poco probable que los peces y otros organismos acuáticos pudieran sobrevivir en un entorno así.
En cuanto al enlace o puente hidrógeno intramolecular, es el que se forma dentro de la misma molécula, estando el hidrógeno, a no más de 2 Ä. Ejemplo:

Como se ve en estos ejemplos, la influencia en la acidez del puente hidrógeno, es notoria, facilitando la expulsión del hidrógeno ácido.
El agua y los enlaces de hidrógeno
Quizás el ejemplo más relevante al que se puede hacer referencia cuando se habla de enlaces de hidrógeno, es el agua. La naturaleza de sus enlaces de hidrógeno le confieren una gran afinidad hacía un sin número de sistemas biológicos, principalmente con las estructuras de las cuales se origina la vida: proteínas, ácidos nucleicos, lípidos y complejos carbohidratados. Todas estas estructuras están directamente relacionadas a su entorno acuoso por sus interacciones inter- e intra-moleculares.
Estudios de RMN-1H en el agua muestran un desplazamiento químico para los protones involucrados en el enlace de hidrógeno a bajas frecuencias, es decir alrededor de +5,5 ppm a campo bajo en fase de vapor a 100 ºC, de modo que, ocurre una disminución en la fuerza del enlace de hidrógeno; mientras que los experimentos de RMN-17O, se observa a campo bajo, en fase gaseosa, la señal producto del oxígeno de la molécula de agua, cuyos valores se encuentran entre +36-38 ppm. Esto evidencia que el enlace de hidrógeno en los dímeros de agua es sustancialmente fuerte, dado que existe aún en el estado gaseoso en concentraciones significativas. No obstante, como las moléculas de agua están relativamente separadas en solución, existe espacio suficiente para que éstas puedan adoptar la configuración que tenga el menor requerimiento energético. Esto lleva a pensar que el enlace de hidrógeno en la molécula de agua tiene consigo un conjunto de movimientos aleatorios con estiramientos y flexiones de todos los enlaces involucrados.
En el agua líquida los arreglos instantáneos de los puentes de hidrógeno no son simétricos. De hecho, estos arreglos son aleatorios, existiendo igual probabilidad de que los cuatro enlaces de hidrógeno (los dos hidrógenos aceptores y los dos hidrógenos donadores) se localicen en cualquiera de los cuatro vértices alrededor del oxigeno. Además, las moléculas de agua rodeadas por los cuatros enlaces de hidrógeno tienden a agruparse en cúmulos, siendo estos favorecidos tanto por razones energéticas como probabilísticas. El efecto cooperativo, observado en las cadenas H-O···H-O···O influye marcadamente en la fuerza del enlace de hidrógeno, encontrándose que en el agua líquida ésta puede llegar a ser 250% mayor que en un dímero de agua.
Las estructuras del hielo, como ya se ha nombrado, proveen los ejemplos más variados y flexibles en relación a la formación del enlace de hidrógeno. Esto se puede derivar de los diferentes polimorfos o fases cristalinas en las que la molécula de agua cristaliza en el hielo. Tales fases cristalinas se conocen como: I, Ih, Ic, II, III, IV, VII, VIII y XII. En el estado sólido la molécula de agua tiene un arreglo inusualmente abierta en sus estructuras cristalinas, producto de la disposición de la molécula de agua en un entorno tetraédrico circundado por las cuatro moléculas de agua más cercanas enlazadas por enlaces de hidrógeno; además, por cálculos de dinámica molecular se demuestra que el agua es capaz de desarrollar 90 configuraciones posibles de celdas unitarias distintas. Una consecuencia notable de la estructura abierta del hielo se refleja en su baja densidad a temperaturas debajo del 0º, que es de 0,92 g mL-1 a diferencia de 1,00 g mL-1 a una temperatura de 0º, y sus consecuencias en la vida de los lagos, ríos y mares.
Importancia del agua y sus interacciones en los sistemas biológicos.
El agua es un componente fundamental en los procesos de vida. No existe especie animal o vegetal que pueda prescindir de la ausencia total del agua. Esto se observa tanto a niveles macroscópicos como microscópicos. Tal es así que cuando se retira el agua de macromoléculas, tales como proteínas y ácidos nucleicos, éstas pierden su función biológica como consecuencia de la degradación de su arquitectura tridimensional. Desde el punto de vista del enlace de hidrógeno, la molécula de agua es la única en poseer doble grupo donador y doble grupo aceptor de enlaces de hidrógeno, los cuales están determinados parcial o totalmente por la combinación de sus propiedades dinámicas de flip-flop y las de cooperatividad de los grupos hidroxilo, que son los responsables de la esfera de hidratación alrededor de las proteínas y los ácidos nucleicos, además de mantener la estructura tridimensional de los mismos, ya sea en su estado natural o cristalino. Por ejemplo, cuando se sintetiza in vitro ribonucleasa A, ésta desarrolla pliegues por la formación de cuatro enlaces de bisulfuro. El acercamiento de estos grupos funcionales contenidos en residuos de cisteína resulta de la asociación cooperativa entre los enlaces de hidrógeno formados por las moléculas de agua con aminoácidos de diferentes dominios de la proteína. Las proteínas globulares de tamaño pequeño contienen moléculas de agua escondidas. Tal es el caso de la proteína inhibidora de la tripsina pancreática. Esta proteína contiene 18 residuos de aminoácidos y 4 moléculas de agua. Otros ejemplos son la lisozima con 129 residuos de aminoácidos y 4 moléculas de agua, y la actinidina con 218 residuos de aminoácidos y con un número de moléculas internas de agua entre 10-20. Estas moléculas de agua pueden permanecer ocultas en el interior de la proteína globular o localizarse en las cercanías de la superficie.
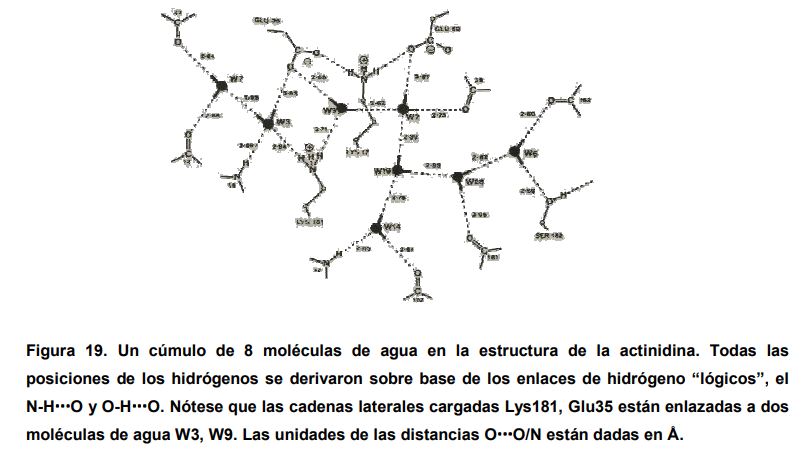
En algunas proteínas emparentadas, las moléculas internas de agua se ubican en las mismas posiciones, por lo que estas moléculas pasan a ser parte indispensable de la estructura tridimensional. Así como sucede con tres cistein-proteinazas vegetales, la actinidina, la papaina y la calotropina D1, donde hay de 15 a 16 moléculas de agua en las mismas posiciones, aproximadamente. En la siguiente figura, se muestra el patrón de enlaces de hidrógeno para estas proteínas que contienen de 8 a 16 moléculas internas de agua.
Las moléculas de agua no sólo se ubican en las cavidades de la estructura proteica, sino que son necesarias para estabilizar el plegamiento tridimensional, cuya función es llevada a cabo por los enlaces de hidrógeno; de lo contrario, se verían poco satisfechos los grupos donadores y aceptores de hidrógeno, además de los grupos medianamente cargados. Por ejemplo, en la penicilopepsina hay varios grupos, 7 grupos C=O y 9 N-H en la cadena principal y 2 cargados en la cadena lateral, donde hay patrones de enlaces de hidrógeno intramoleculares, y en la lisozima sucedería lo mismo con 3 grupos C=O y 3 N-H internos en la cadena principal si no hubiese moléculas de agua internas.
En algunos casos, estas moléculas de agua forman enlaces de hidrógeno con los grupos cargados de las cadenas laterales, lo cual previene el colapso estructural de los residuos de aminoácidos cargados de la cadena opuesta. Esto se observa en la carboxipeptidasa entre la Asp104-agua-Asp59 y en la actinidina en los residuos Glu35- agua-Lys181. Tales moléculas de agua están rodeadas, en gran parte, por un ambiente apolar, donde sus enlaces de hidrógeno con frecuencia son fuertes y están bien definidos. En las estructuras cristalinas de moléculas medianas de proteínas, las moléculas de agua están unidas, comúnmente, a tres o cuatro enlaces de hidrógeno simultáneamente, con una participación de las interacciones del 48% para los tres enlaces y 37% para los de cuatro. Además, las moléculas de agua en la estructura de las proteínas no se encuentran solas sino que están ensambladas en forma de cúmulos. En algunas de la estructura de proteínas, una molécula de agua interna se asocia con tres grupos aceptores indicados en los enlaces triple-centrados como los observados en las estructuras de cristalinas de los α-aminoácidos zwitteriónicos.
Otro ejemplo de la importancia del agua en la funcionalidad de las proteínas lo representa el control que ésta ejerce en el flujo de los iones sodio, calcio y potasio a través de los canales inter- y extra-membranales de las células en la regulación de las señales nerviosas. También existen en las membranas celulares los denominados canales de agua (ver figura) que facilitan el rápido transporte de agua en respuesta a un gradiente osmótico. En las proteínas, las moléculas de agua no sólo hidratan la superficie de éstas, sino que también ocupan los intersticios entre moléculas de la proteína en la red cristalina, encontrándose en posiciones específicas. Además, las interacciones con los átomos o grupos de átomos de la cadena lateral y principal merecen especial atención.

Los aminoácidos y los enlaces de hidrogeno
Definición y naturaleza
En principio, el término aminoácido se podría referir a cualquier compuesto que tenga una función amino y otra acídica. Sin embargo, en la práctica, la palabra aminoácido ha sido principalmente empleada para hacer referencia a la α-aminoácido proveniente de fuentes naturales. Se han aislado e identificado 100 de estos aminoácidos, pero sólo 20 de ellos se obtienen de la hidrólisis de las proteínas más comunes. Los 20 α-aminoácido que conforman los bloques estructurales de las proteínas se muestran en la Tabla I.6. Todos ellos tienen grupos aminos primarios. Todos ellos, excepto la glicina, tienen al menos un átomo de carbono asimétricamente sustituido, el carbono α, por lo que se encuentran en la naturaleza en su forma óptimamente activa.
Los aminoácidos son sólidos muy solubles en agua. Esta propiedad se mantiene en todos los aminoácidos, inclusive en los β-, γ-, δ-, ε-aminoácidos (refiriéndose a la posición del grupo amino con respecto al grupo carboxilato), pero va disminuyendo en la medida que las cadenas alifáticas se van haciendo más largas. Esta tendencia se observa claramente en la Tabla I.6, donde la solubilidad disminuye gradualmente desde la glicina hasta la leucina a medida que aumenta el tamaño del grupo R. Además, los aminoácidos presentan elevados puntos de fusión si se comparan con moléculas orgánicas de tamaño y peso molecular similar. Esto sugiere que la estructura de estos compuestos debe ser semejante a la de una sal y, por lo tanto, la misma puede escribirse como la de un ion híbrido, llamado zwitterión, tal y como se muestra en la Figura 24.



Por otra parte, la estructura molecular de los aminoácidos está determinada, en parte, por el pH del medio. Por ejemplo, a valores de pH<1 la glicina existe como un catión protonado A, pero a pH>12 el anión aminocarboxilato C es la forma principal. Entre estos dos extremos en pH, la sal interna B se presenta en equilibrio entre las estructuras A y C.

Por consiguiente, los valores de pH en los aminoácidos se asemejan mucho a los de un ácido orgánico biprótico. La curva de titulación con una base acuosa indica que, aproximadamente, el pKa1 para el ácido carboxílico tiene un valor de 2,3 y el pKa2 para el ión amonio es cercano a 9,5. El pH para el cual un aminoácido es una especie eléctricamente neutra se conoce como punto isoeléctrico, pI o pHI. Este valor en la mayoría de los aminoácidos se encuentra entre la mitad de los valores de pKa1 y pKa2,
pI = ½ (pKa1 + pKa2)
En la Tabla 7, se observan los valores de los pKa y pI para los aminoácidos fundamentales.

El grupo carboxílico de un α-aminoácido protonado es más acídico que aquellos ácidos monocarboxílicos en al menos dos unidades de pKa. Esta propiedad es atribuida a la atracción de electrones por efecto inductivo del grupo amonio adyacente. El valor de pKa1 de un aminoácido se incrementa si el grupo amonio se mueve más lejos del grupo carboxílico, es decir;

Esta tendencia se puede observar más claramente en la Tabla 8, donde la acidez de los grupos carboxílicos está estrechamente relacionada con la separación de los grupos en una cadena carbonada cíclica o acíclica.

El enlace de hidrógeno en los aminoácidos
El enlace de hidrógeno en los α-aminoácidos está dominado por los grupos amonio y carboxilato. A estos grupos se les atribuye el 52% de los enlaces de hidrógeno formados, y el 48% restante se distribuye entre los grupos O-H (24%), N-H (11%) y C-H (13%). Por otra parte, el 80% de los enlaces de hidrógenos con los grupos – +NH3 forman enlaces bifurcados. Esto refleja la disparidad entre las propiedades del grupo – +NH3, que provee sólo tres protones para formar enlaces de hidrógeno, y el grupo -COO- , que es capaz de aceptar seis enlaces de hidrógeno. El enlace de hidrógeno en los aminoácidos es difícil de definir; sin embargo, en ellos se pueden deducir cuatro diferentes grupos donadores, el +N-H, N-H, O-H y C-H, y nueve grupos aceptores el -COO- , -COOH, -O-H, OH2, Cl- , -OSO3 – , –NH2, –NH y =N. El grupo donador de enlaces de hidrógeno predominante en los aminoácidos es el –+NH3 y el aceptor es carboxilato –COO- . Por otra parte, sólo los grupos hidróxilo y el nitrógeno imidazólico tienen las propiedades potenciales para ser grupo aceptor y donador.
El enlace de hidrógeno en la estructura cristalina de los aminoácidos se rige más o menos por las siguientes reglas:
- La energía máxima de contribución del enlace de hidrógeno a la energía reticular está dada por un número igual de grupos aceptores como donadores. El enlace de hidrógeno en la red cristalina de los aminoácidos es uno de los factores principales que determina su empaquetamiento molecular. Además, en el empaquetamiento cristalino también pueden influir la segregación de los grupos polares y los no polares.
- El enlace de hidrógeno está representado, principalmente, por el enlace zwitteriónico +N-H···OCO. Esto se muestra por una simple inspección de los patrones de enlaces de hidrógeno, donde se necesitan satisfacer las propiedades inherentes a cada uno de estos grupos. El esquema de enlaces de hidrógeno que se puede adoptar en un determinado empaquetamiento cristalino dependerá de cada molécula de aminoácido. El grupo donador tiene un número invariable de tres enlaces de hidrógeno, mientras que cada átomo de oxígeno del grupo aceptor puede aceptar al menos dos enlaces, aunque frecuentemente acepte tres. La capacidad aceptora de un grupo carboxilato nunca es menor de cuatro y puede ser tan grande como un número de siete enlaces. Los grupos cetónicos y los ácidos carboxílicos presentan una demanda menor y aceptarán un solo enlace, aunque, dos es lo más común.
- Los enlaces de hidrógeno N-H…O más fuertes se dan entre el N-H de un anillo imidazólico y el oxígeno de un grupo carboxilato; y el enlace O-H…O más fuerte se da entre el O-H del ácido carboxílico y el átomo de oxígeno de una molécula de agua, del grupo carboxilato o del grupo sulfato. Estos se forman siempre y cuando estos grupos estén presentes en las moléculas, y en el caso del agua, co-cristalicen con ellas.
- Existen evidencias claras de los enlaces de hidrógeno C-H…O en las estructuras cristalinas de aminoácidos. Estos son más comunes entre los carbonos α y oxígenos de los grupos carboxilato.
Enlace de hidrógeno bifurcado donador en aminoácidos
El papel de los enlaces bifurcados donadores es el de poder proporcionar todos los medios para incluir a los átomos aceptores de enlaces de hidrógeno en el esquema de enlaces de hidrógeno cuando haya deficiencias de protones. Es importante, notar que las fuertes propiedades aceptoras de los átomos de oxígeno de los grupos carboxilato y carbonilo parecen dominar los patrones de enlaces de hidrógeno en los aminoácidos, en comparación a las propiedades direccionales de los enlaces donadores. Esta definición proviene de una comparación detallada de la geometría del enlace +N-H…O en los aminoácidos. Esta observación es consistente con el concepto de enlace de hidrógeno como una combinación de fuerzas coulómbicas y de intercambio, las cuales son diferentes por las distintas combinaciones de los grupos donadores y aceptores. Desde luego, el enlace de hidrógeno bifurcado donador es difícil de definir en los aminoácidos, ya que existe un rango continuo donde los enlaces dobles centrados y bifurcados donador no son diferenciables. No obstante, existe un ángulo +N-H…O y una distancia de enlace H…O de ~130º y ~2,1Å, respectivamente. Por consiguiente, se puede adoptar la siguiente definición empírica para el enlace bifurcado donador, dada en la Figura 25.

En esta geometría, el átomo de hidrógeno se encuentra muy cerca del plano X, A1,A2, y está atraído por los átomos A2 y A1, con una distancia de enlace de hidrógeno normal H…A1, d1, de 1,4 Å, siendo A1 el único átomo electronegativo dentro de un radio de 2,85Å. Se puede considerar que la distancia más corta, d1, corresponde a la del enlace de hidrógeno, mientras que la más larga, d1, es una perturbación electroestática. En la Tabla 9, se dan los valores de los parámetros geométricos θ1, θ2, θ3, d1 y d2 obtenidos por difracción de neutrones para algunas estructuras cristalinas de aminoácidos.

Efectos cooperativos y patrones de enlaces de hidrógeno en los aminoácidos.
Los patrones de enlaces de hidrógeno en las estructuras cristalinas de aminoácidos están sujetos a efectos cooperativos. El efecto cooperativo en los enlaces de hidrógeno puede formar estructuras extendidas, que en el caso particular de los cristales, forman arreglos periódicos moleculares de carácter bi- y tri-dimensional. Debido a la polarizabilidad o a la transferencia de carga de estas estructuras extendidas construidas con n moléculas unidas por enlaces de hidrógeno, la energía del enlace de hidrógeno en esas estructuras es mayor que la suma de los enlaces individuales, es decir;
E(H···A)n > nE(H···A).
En consecuencia, la cooperatividad es no-aditiva. Los primeros estudios de cooperatividad en los enlaces de hidrógeno fueron realizados en agua líquida y soluciones acuosas. Estudios sistemáticos en estructuras cristalinas de carbohidratos demostraron que la cooperatividad produce un acortamiento en la longitud de los enlaces de hidrógeno, y en el caso de trímeros de metanol produce una ganancia de energía por enlace de hidrógeno de 12%, aproximadamente. Por otra parte, las interacciones cooperativas son más importante en la fase sólida que en las fases gaseosa o líquida, porque las interacciones débiles entre moléculas vecinas en la estructura cristalina pueden desarrollarse más efectivamente en sistemas ordenados. Finalmente, los efectos cooperativos pueden jugar un papel preponderante en la determinación de estructuras de materiales construidos a partir de unidades moleculares discretas, como es el caso de los cristales moleculares, y en las estructuras supramolecular en polipéptidos. Un ejemplo donde se puede apreciar los efectos cooperativos lo representa la estructura de la glicina, en la cual se observa la formación de doble cadenas infinitas de aminoácidos, en las cuales las moléculas se alinean e interactúan de manera cabeza-cola, es decir a través de enlaces … +N-H…OCO- …; adicionalmente, las moléculas interactúan lateralmente, también por enlaces de hidrógeno, para formar cinta infinitamente extendidas, así como se observa en la Figura 26.

